
23.09.11 Граница этапов эпохи Плиоцена (возраст 2,6 млн. лет) является датой мутаций, обнаруженных у всех современных людей. Эти мутации предшествуют времени возникновения принципиально нового вида Номо еrectus [Wood, 2002].
Этот вид отличается от всех ранее живущих гоминид резким возрастанием
объема головного мозга и считается предком современного человека [Hawks, et al., 2000].
Мутация, в результате которой был инактивирован ген, кодирующий
наработку сахарозы на поверхности клетки, имеет возраст ~ 2,8 млн. лет [Chou, et al., 2002].
Примерно этот же возраст (~ 2,4 млн. лет) имеет мутация гена миозина, в
результате которой произошло значительное уменьшение объема и силы
мощных жевательных мускулов, характерных для человекообразных обезьян и,
по словам авторов [Stedman, et al., 2004],
были ликвидированы анатомические ограничения роста объема черепной
коробки, и, как следствие, увеличения объема головного мозга.
К
событиям этого времени (2,60 +/-0,94 млн. лет) следует отнести недавно
обнаруженное разделение африканских слонов на два вида [Roca, et al., 2001].
Заметим, что предшествующее этому разделение слонов на азиатских и
африканских, по времени совпадает с расхождением генетических линий
человека и шимпанзе (~ 5 млн. лет) [Roca, et al., 2001].
Сравнение
мтДНК современных людей и мтДНК, выделенной из останков неандертальцев,
до недавнего времени считавшихся нашими предками [Rozzi, Castro, 2004] показало, что их линии являются параллельными, а не последовательными [Serre, et al., 2004]. Разъединение между линиями человека и неандертальца произошло около 500 тыс. лет назад [Takahata N., Lee S. and Satta Y. 2001], что также находит отражение в палеомагнитной шкале.
Возраст общего предка современных людей и место его возникновения оценивался различными авторами [Cann, et al., 1987; Vigilant, et al., 1991; Horai, et al., 1995; Takahata, et al., 2001].
По последним оценкам средний возраст самой древней обнаруженной мтДНК
составляет 230 тыс. лет, и 100 тыс. лет – для У-хромосомы, причем, оба
имеют африканские корни [Cavalli-Sforza L., Feldman M., 2003].
Однако это совсем не означает, что в эти периоды существовало по одной
женщине, или по одному мужчине, или не было линий-современников мтДНК и
У-хромосомы. Считается, что к нашему времени сохранились потомки только
этих линий. Дата возникновения этих мтДНК и У-хромосомы соответствует
событиям, отраженным на палеомагнитной шкале, а именно моментам
экскурсов геомагнитного поля.
Филогенетическая
схема, составленная антропологами исходя из найденных останков, дает
представление о временных отрезках существования разных видов гоминид.
По этой схеме можно видеть, что этап от 5 до 2 млн. лет соответствует
гоминидам с маленьким объемом головного мозга [Takahata, et al., 2001], не менявшимся в течение этого периода. Обнаружено, что увеличение размеров мозга у представителей семейства [Wood, 2002] произошло в эпоху Плейстоцена. Последние находки новых видов гоминидов заставляют авторов [Wood, 2002]
отойти от названия представленной схемы «деревом», согласно которой
представленные виды связывались в последовательные цепочки
«предок-потомок». Целесообразность нового подхода подтверждается
антропологическими характеристиками последних находок останков гоминид,
схожих с представителями более позднего времени [Leakey, et al., 2001; Brunet, et al., 2002]. Антропологи [Wood, 2002]
склонны представлять эволюцию гоминид сродни разрастанию куста, когда в
течение данного периода появляется сразу несколько видов гоминид,
причем причина такого разрастания не называется.
 Таблица 2. Этапы эволюции человека, палеонтологическая и палеомагнитная шкалы
Возникновение рас (географических популяций) и схема расселения
Определение расы звучит примерно так: «раса – это большая популяция индивидов, у которых значительная часть генов общая и которую можно отличить от других рас по общему для нее генофонду». На основании числа аллелей, свойственных той или иной группе организмов, возможно определение генетического расстояния между ними. Оказалось, что для больших рас эта величина выражается числом 0,03. Это гораздо ниже значений, характеризующих подвиды (0,17–0,22) и тем более виды животных (0,5–0,6 и более). В животном мире генетическое расстояние, равное 0,03, соответствует отличиям местных популяций друг от друга.
Различия между расами объясняется естественным отбором. Принято считать, что естественный отбор является главным фактором эволюции внешних признаков (фенотипов). Он обуславливает приспособление к различным условиям окружающей среды. Однако чтобы действие отбора закрепилось, необходимо наличие репродуктивной изоляции. Существовал ли исторический период, когда все человечество было разделено на три более или менее изолированные группы?
На протяжении большей части последнего ледникового периода (около 100 тыс. лет назад) громадная площадь Земли была покрыта льдом. Гималаи и Алтайские горы с расположенными на них ледниками разделили Евроазиатский континент на три области, создавая тем самым условия для раздельной эволюции белых на западе, монголоидов на востоке и негроидов на юге. Современные области расселения трех больших рас не совпадают с теми областями, в которых они формировались, но это можно объяснить более поздними миграциями. По каким же признакам шел отбор? Скорее всего, это были приспособления к конкретным природно-климатическим условиям.
Традиционно выделяют три основные расы: европеоиды (или евразийская раса), негроиды (экваториалы) и монголоиды (азиатско-американская раса). Но многие антропологи считают, что с биологической точки зрения рас значительно больше – как минимум 8 или 10. В
частности, можно назвать южноафриканскую (бушмены и готтентоты),
австралоидную, айноидную, американоидную расу и ряд других. Их
представители отличаются по некоторым существенным морфологическим
признакам, таким как цвет кожи, глаз и волос, особенности строения лица и
т.д. Существуют чисто биологические механизмы разделения на расы.
Принято считать, (во-первых) для того, чтобы образовалась группа со
своеобразным генофондом, необходима изоляция – тогда в силу принципа
случайности мутаций (как по конкретному гену, так и по времени
возникновения) группа начинает автоматически дивергировать, чему
способствует также вероятностный характер закрепления новых мутаций.
Считается так же (во-вторых), что в различных климатогеографических
зонах в ходе адаптации и естественного отбора возникают признаки,
способствующие выживаемости в данной области. Рассмотрим этот вопрос в
контексте нашей модели, для чего обратимся к схеме возникновения
человеческих рас (рис. 8).
 Рис. 8. Схема возникновения человеческих рас [Cavalli-Sforza, Feldman, 2003].
На
схеме показано расхождение между расами в генетических расстояниях.
Видно, что африканская раса не изменялась на дистанции 0.205, в то время
как европейцы и североафриканцы подверглись трем бифуркациям. Последняя
бифуркация соответствует дистанции 0.015. Генетические расстояния (ГР)
определяются по частоте рекомбинаций между генетическими маркерами. Это
количественная характеристика сравниваемых объектов (популяций, видов и
т.п.), определяющая среднее число замен нуклеотидов в каждом локусе,
которые произошли за время раздельной эволюции двух групп организмов; в
настоящее время наиболее широко применяется метод расчета ГР по Нэю (Nei genetic distance).
Иногда ГР указывается в морганидах (сантиморганидах, сМ), иногда, – во
временных интервалах (в млн. лет) между бифуркациями. На схеме выделены
ГР: ~ 200 тыс. лет, ~100 тыс. лет, 46 и 15 тыс. лет. Эти даты – периоды
прохождения соответствующих экскурсов эпохи Брюнес. Если совпадения не
случайны и ГР действительно выражены во временных интервалах (в млн.
лет), то разделение человечества на расы так же связано с повышением
радиационного фона на Земле в периоды экскурсов (см. Табл. 2).
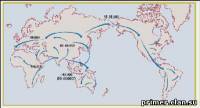 Рис. 9. Карта расселений [Cavalli-Sforza, Feldman, 2003].
Карта расселений, непосредственно связанных с возникновением рас, показана на рис. 9. Человечество
начало расселяться по материкам из Африки примерно 100 тыс. лет тому
назад. Анализируя пути и сроки миграций, отметим две интересные детали.
Первая касается заселения Австралии примерно 50-60 тыс. лет тому назад.
Как следует из модели горячей Земли [Кузнецов, 1990],
в это время в Австралию можно было перебраться практически по суше.
Вторая деталь, аналогичная первой, относится к переселению наших предков
через Берингов пролив, который, как сейчас известно, образовался
сравнительно недавно, во время т.н. «Великого потопа», в момент таяния
ледников сразу после экскурса Гетеборг, 15 тыс. лет тому назад. Именно
поэтому последний срок переселения предков на Американский материк
датируется временем этого экскурса. Почему Африка? Один
из принципиальных вопросов из области проблем эволюции человека
заключается в том, почему местом рождения современного человека является
Африка? Почему все попытки найти следы возникновения (но не миграции)
гомо сапиенс на других континентах оканчивались безрезультатно? Почему
первобытные люди стали расселяться из Африки? Почему именно в Африке
находят следы генетических мутаций многих других биологических видов? Ответом на эти вопросы было бы доказательство того, что именно в Африке радиационный фон в период инверсий и экскурсов был наибольшим, значительно превышающим фон на других континентах.
Обратимся к опубликованным данным по поведению геомагнитного поля в
периоды одних и тех же инверсий и экскурсов, записанных в осадках или
вулканических извержениях, произошедших в различных регионах Земли. Авторы [Hartl, Tauxe, 1996]
показали, что уменьшение модуля поля в момент инверсии Брюнес-Матуяма и
предшествующего инверсии экскурса Хурамильо, было различным для
различных регионов. Пробы брались в районах расположения базальтов,
излившихся на дно океанов вблизи Индонезии (0., 160.E) и западного
берега Африки (0., 20.W). В первом случае уменьшение модуля поля не
превышало 1/10 от его нормальной величины, во втором, - понижение модуля
было большим, не менее 1/30 от нормального значения. Эти данные
подтверждаются другими аналогичными наблюдениями. Так, например, в [Oda, et al., 2000]
показано, что в момент этой инверсии в базальтах, излившихся в районе
острова Барнео (7. N, 122.E), модуль поля уменьшился примерно в 5-7 раз
от его нормальной величины. Аналогичное по величине уменьшение было
зафиксировано вблизи побережья Калифорнии [Heider, et al., 2000]. Значительное понижение модуля было зарегистрировано вблизи западного побережья Африки [Yamazaki, Oda, 2001].
Авторы зарегистрировали уменьшение модуля геомагнитного поля в периоды
инверсии Брюнес-Матуяма и экскурса Хурамильо, не менее чем в сто раз. Приведенные
данные по поведению модуля поля в моменты инверсий можно считать
подтверждением теории дрейфа магнитных полюсов в моменты инверсий и
экскурсов. Согласно этой теории дрейф полюсов происходит по вполне
определенным траекториям, на которых располагаются глобальные магнитные
аномалии [Kuznetsov, 1999].
В момент времени инверсии (экскурса), когда поле дипольного источника
становится близким к нулю, собственное поле глобальных магнитных
аномалий ещё сохраняет свое прежнее значение. Поле дипольного источника
возрастает от нулевого значения со знаком, обратным предыдущему. Это
приводит к аннигиляции поля дипольного источника и поля аномалии. Так
происходит смена полярности поля на выделенных трассах. В тех районах,
где влияние аномалий не сказывается, например, в Африке, модуль поля во
время инверсий и экскурсов становится практически равным нулю. Таким
образом, палеомагнитные данные однозначно показывают, что в момент
инверсии, – модуль геомагнитного поля в Африке уменьшается значительно
больше (в 100 и более раз), по сравнению с другими регионами (Азией,
Австралией, Америкой), в которых модуль поля уменьшается не более чем в
10 раз (рис. 10, 11).

Рис. 10. Уменьшение интенсивности геомагнитного поля в период инверсии Брюнес-Матуяма
(DIP 2) и предшествующего инверсии экскурса Хурамильо (DIP 1) [Hartl,
Tauxe, 1996]. Черными точками и линиями показаны места взятия проб.
 Рис. 11. Изменение относительной палеоинтенсивности геомагнитного поля в период инверсии Брюнес-Матуяма, по данным образцов [Yamazaki, Oda, 2001], взятых на западном побережье Африки (20º S, 10º E).
Рост уровня радиации при уменьшении модуля геомагнитного поля
Построим
ориентировочную, феноменологическую зависимость радиационной дозы D от
величины модуля геомагнитного поля В. Очевидно, что в момент инверсии
(экскурса), когда модуль поля значительно уменьшается, величина
корпускулярного потока, попадающего на Землю, возрастает, иначе,
увеличивается уровень радиации Земли. В целях упрощения оценок, будем
полагать, что приходящая на Землю радиация (в зивертах) биологическими
объектами, обитающими на Земле, полностью поглощается, при этом
поглощенная доза (в греях) точно равна уровню радиации (в зивертах).
Такое допущение, в частности, означает, что радиационный фон ГКЛ*
(равный на поверхности Земли ~ 0.5×10-3 Зв) эквивалентен дозе (~ 0.5×10-3 Гр). Основанием к такому допущению служит то обстоятельство, что один зиверт, и один грей, – оба равны 104 эрг/г.
При
построении зависимости: «радиационная доза – модуль геомагнитного поля»
учтем величины общепринятых значений дозы. Будем считать, приближенно,
что при уменьшении модуля геомагнитного поля в 100 раз, радиационная
доза возрастает, по крайней мере, не менее чем в 100 раз. В
действительности, увеличение дозы может быть даже больше. Докажем это
утверждение. Энергия частиц, отклоняемых геомагнитным полем Е примерно
пропорциональна величине геомагнитного поля В: Е ~ В. На заряженную
частицу, попадающую в магнитное поле, как известно, действует сила
Лоренца F = Z evB, равная центробежной силе mv2/R, иначе: mv =
Z evR. Переходя к более удобным единицам, получаем: E = 300 ZBR, где: E
– энергия частицы в МэВ, Z – заряд частицы, B – индукция магнитного
поля в Тл, R – радиус кривизны траектории частицы в м, е – заряд
электрона, v – скорость частицы, m – её масса.
Принято
считать поток космических лучей (КЛ) величиной фрактальной. Это
означает, что число частиц потока КЛ N, обладающих энергией Е: N(E) ~ Е-d,
где 1 < d < 2, d – фрактальная размерность. Учет этого
обстоятельства позволяет ориентировочно оценить, как будет возрастать
радиационная доза, при уменьшении модуля геомагнитного поля. Например,
если модуль поля уменьшится в период инверсии в 100 раз, то доза может
увеличиться даже больше чем в 100 раз, в пределе, в 10 000 раз. Докажем
возможность такой оценки. Как было показано, чем меньше величина
магнитной индукции, тем частицы меньшей энергии способны преодолевать
защитную магнитную оболочку Земли: Е ~ В. Это соответствует тому, что
плотность потока КЛ, попадающего на Землю, возрастает. Доза
ионизирующего излучения на поверхности Земли D определяется плотностью
потока частиц Р, коэффициентом передачи энергии L и временем действия
ионизирующего излучения t: D = PLt. Очевидно, что если параметры Lt – не
меняются, то плотность потока Р ~ N(Е), следовательно, и D ~ N(Е). Учитывая принятое нами ранее N(E) ~ Е-d, где 1 < d < 2, то и доза D ~ В-d.
Т.о. мы получили зависимость, показывающую, что уменьшение величины
геомагнитного поля приводит к росту дозы ионизирующего излучения. Речь
идет пока только об эффекте уменьшения отклонения заряженных частиц при
уменьшении величины магнитной индукции Земли. Если учесть, что при этом
будут разрушаться радиационные пояса, то поток заряженных частиц на
Землю возрастет ещё больше, что приведет к ещё большему возрастанию дозы
ионизирующего излучения.
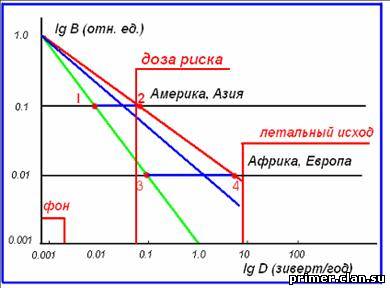
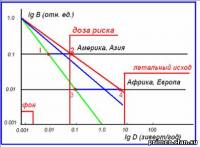 Рис.12. Зависимость радиационной дозы (D) от величины модуля геомагнитного поля (B).
Фрактальная
размерность d, согласно нашей модели, может меняться от 1 до 2. При
этом радиационная доза D, в зависимости от величины В, может меняться от
D ~ В (линия 1 – 3 на рис. 16) до D ~ В-2 (линия 2 – 4). На
этом рисунке, на шкале lg D (в зивертах/год) отмечены три характерных
значения дозы: фон ≈ 2 - 3 мЗв; доза риска ≈ 60 мЗв; доза летального
исхода ≈ 6 - 8 зиверт/год.
Сравним
величину дозы, приходящейся на два различных региона, в одном из
которых модуль геомагнитного поля в период инверсии уменьшился в 10 раз
(Америка, Азия), с другим, в котором поле уменьшилось в 100 раз.
Предположим, что величина фрактальной размерности d ≈ 1.4. Надо сказать,
что такое значение величины d характерно для природных процессов,
имеющих фрактальный характер (синяя линия на рис. 12). Будем
рассматривать область значений, при которых радиационная доза больше
дозы риска. При этом оказывается, что в Америке и Европе доза риска
(начало генетических мутаций) вообще не будет достигнута, в то время как
в Африке доза может быть порядка зиверт/год. Известно, что при дозе,
большей, чем 200 мЗв/год, преобладает массовая гибель клеток.
Приведенные оценки показывают, что радиационная доза может сильно
отличаться для различных регионов мира при едином, общем для Земли,
явлении инверсии или экскурса.
Заключение
В
течение последних 10 – 20 лет учеными получены новые данные по
изменениям климата и эволюции животного мира и, в значительной степени, –
эволюции человека, происходивших на протяжении сотен тысяч и миллионов
лет, предшествующих современному периоду. Полученные новые данные, как
правило, находились в противоречии старым, привычным и устоявшимся
догмам.
Повторяемость
оледенений на Земле с примерной периодичностью в 100 тыс. лет
стимулировала, в своё время, создание теории вариаций климата
Миланковича, согласно которой климатические изменения на Земле являются
откликом на вариации в её орбите. Причина вариаций, происходящих с
основными периодами 19, 23 и 41 тыс. лет, связана с возмущениями орбиты
Земли за счет её эксцентриситета (периоды: 95, 136 и 413 тыс. лет),
прецессии (19 и 23 тыс. лет) и наклона плоскости эклиптики. Из теории
Миланковича следует, в частности, что изменения климата на Земле должны
происходить в её северном и южном полушариях – в противофазе. При
изучении кернов льда, извлеченных в Антарктиде и Гренландии, показано
практически полное совпадение данных по синхронному изменению климата в
обоих полушариях. Имеется достаточно много других данных, противоречащих
теории Миланковича [Большаков, 2001].
Результаты,
достигнутые в антропологии и генетике, так же как и в случае с
климатом, оказываются несовместимыми с представлениями об эволюции жизни
на Земле вообще и эволюции человека, в частности. Генетики имеют веские
доказательства того, что не существует единого генетического «дерева»,
на котором можно было бы найти «ветку», ответственную за современного
человека и, более того, определить другую «ветку», на которой выросла
наша, человеческая. Сенсационными оказались открытия генетиков по поводу
обнаружения «Африканской Евы» – нашей прародительницы. в то же время
такой вопрос, как разделение
человечества на расы, всё ещё объясняется замкнутостью популяции. Хотя
каждому и так очевидно, что, сколь долго ни изолируй европейцев в
Африке, они всё равно не станут неграми.
В настоящей работе показано, что экскурсы
геомагнитного поля совпадают с периодами резкого потепления климата и
исчезновения пылевой компоненты в антарктическом и гренландском льдах.
Как было показано в работе, эти совпадения не случайны. Наличие
экскурсов является причиной изменений, происходящих на поверхности
Земли. Можно предположить, что в периоды, соответствующие увеличенной
концентрации пыли, на Земле было не только холодно, но и не видно
Солнца, и жизнь на Земле, надо думать, не была комфортной. Радиационный
фон на поверхности Земли в периоды оледенений был низким и, по-видимому,
не было причины для генетических мутаций. Эволюция биоты происходила
крайне медленно. Но вот, в ядре Земли происходит смена режима работы
фазового перехода. С изменением направления фазового перехода меняется
полярность геомагнитного поля. Земля на некоторое время теряет свою
защитную оболочку, радиация достигает её поверхности, вызывая, с одной
стороны, потепление климата и, с другой, - повышение радиационного фона.
Как следует из нашей модели, потепление климата связано с разрушением
пылевого аэрозольного стратосферного слоя, а проникновение ионизирующего
излучения в атмосферу Земли может значительно (в сотню раз) увеличить
её радиационный фон.
artefact-2007.blogspot.com
|




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}